La herencia genética es el proceso por el cual las características de los individuos se transmiten a su descendencia, ya sean características fisiológicas, morfológicas o bioquímicas de los seres vivos bajo diferentes condiciones ambientales.
La herencia es el proceso por el cual los genotipos crecen y sólo representa una parte de la herencia, es decir, el porcentaje de la variabilidad fenotípica debido a efectos genéticos aditivos. Pero definir las fuentes y el origen de las semejanzas entre miembros de una misma familia incluye también otro tipo de variables. El estudio de la herencia cuantifica la magnitud de la semejanza entre los familiares y representa el porcentaje de variación que se debe a todos los efectos aditivos familiares incluyendo la epidemiología genética aditiva y los efectos del medio ambiente. En los casos en que los miembros de una misma familia conviven resulta imposible discriminar las variables genéticas fenotípicas de las del entorno y medio ambiente. Los estudios de mellizos separados al nacer y de hijos adoptivos permiten realizar estudios epidemiológicos separando los efectos hereditarios entre los de origen genético y las etiologías más complejas, incluyendo las interacciones entre los individuos y la educación. Además, diversos factores influyen en el momento de interpretar los estudios de la herencia incluyendo los supuestos previos por parte de los investigadores.2
Uno de los debates entre los científicos es cual es el peso de la naturaleza y cual es el peso de la cultura, es decir, la magnitud de la influencia de los genes versus la magnitud de la educación y el medio ambiente.3 El dilema de una oposición entre naturaleza y cultura, lo innato versus lo adquirido, es decir innato o adquirido, frase acuñada por Francis Galton en el siglo XIX.
Está comprobado que en los genes se transmite el color de la piel, del cabello, de los ojos. ¿Se transmite también algo de la personalidad, los gustos, el carácter, las capacidades o la inteligencia?
El doctor Albert Rothenberg, profesor de psiquiatría en la Universidad de Harvard y la doctora Grace Wyshak, profesora de psiquiatría en la misma universidad, estudiaron el árbol genealógico 435 de los 488 químicos, físicos, médicos y fisiólogos galardonados con el Premio Nobel entre 1901 y 2003, y el de 50 escritores ganadores del Premio Nobel de literatura y 135 ganadores del Premio Pulitzer. Los resultados de sus investigaciones contradicen la teoría de la transmisión directa del genio del británico Francis Galton publicada en el libro «Hereditary Genius». Según Albert Rothenberg y Grace Wyshak la genialidad no depende de los genes sino de una constelación de factores que no son genéticos sino psicológicos. Los procesos afectivos y cognitivos involucrados en la creatividad son el resultado de una combinatoria de educación, genética y factores sociales. Los premiados no llevaban la genialidad grabada en el ADN ni provenían de familias con coeficientes intelectuales privilegiados sino que se habían educado con el incentivo y la orientación creativa de sus padres. Los deseos incumplidos de estos padres buscaban realizarse a través de sus hijos
Definición de herencia como herencia genética
La herencia genética es la transmisión a través del material genético existente en el núcleo celular, de las características anatómicas, fisiológicas o de otro tipo, de un ser vivo a sus descendientes.
La herencia consiste en la transmisión a su descendencia los caracteres de los ascendentes. El conjunto de todos los caracteres transmisibles, que vienen fijados en los genes, recibe el nombre de genotipo y su manifestación exterior en el aspecto del individuo el de fenotipo. Se llama idiotipo al conjunto de posibilidades de manifestar un carácter que presenta un individuo.
Para que los genes se transmitan a los descendientes es necesaria una reproducción idéntica que dé lugar a una réplica de cada uno de ellos; este fenómeno tiene lugar en la meiosis.
Las variaciones que se producen en el genotipo de un individuo de una determinada especie se denominan variaciones genotípicas. Estas variaciones genotípicas surgen por cambios o mutaciones (espontáneas o inducidas por agentes muta génicos) que pueden ocurrir en el ADN. Las mutaciones que se producen en los genes de las células sexuales pueden transmitirse de una generación a otra. Las variaciones genotípicas entre los individuos de una misma especie tienen como consecuencia la existencia de fenotipos diferentes. Algunas mutaciones producen enfermedades, tales como lafenilcetonuria, galactosemia, anemia falciforme, síndrome de Down, síndrome de Turner, entre otras. Hasta el momento no se ha podido curar una enfermedad genética, pero para algunas patologías se está investigando esta posibilidad mediante la terapia génica.
Lo esencial de la herencia queda establecido en la denominada teoría cromosómica de la herencia, también conocida como teoría cromosómica de Sutton y Boveri:
Los genes están dispuestos linealmente en los cromosomas.
La recombinación de los genes se corresponde con el intercambio de segmentos cromosómicos (Crossing over).
La transferencia genética horizontal es factor de confusión potencial cuando se infiere un árbol filogenético basado en lasecuencia de un gen. Por ejemplo, dadas dos bacterias lejanamente relacionadas que han intercambiado un gen, un árbol filogenético que incluya a ambas especies mostraría que están estrechamente relacionadas puesto que el gen es el mismo, incluso si muchos de otros genes tuvieran una divergencia substancial. Por este motivo, a veces es ideal usar otras informaciones para inferir filogenias más robustas, como la presencia o ausencia de genes o su ordenación, o, más frecuentemente, incluir el abanico de genes más amplio posible.
Críticas a la definición de herencia como herencia genética
La Teoría de los sistemas de desarrollo (DST) se opone a la definición de herencia como transmisión de genes y aplica el concepto a cualquier recurso que se encuentre en generaciones sucesivas y que contribuya a explicar por qué cada generación se parece a la que le precede. Estos recursos incluyen factores celulares y factores externos como la gravedado la luz solar. La DST utiliza, por tanto, el concepto de herencia para explicar la estabilidad de la forma biológica de una generación a otra. La herencia genética es el resultado de la unión de dos células madre que al unirse crearon un fenotipo con características similares de dos, pero sin embargo es un organismo diferente ya que al unirse se crearon nuevas células modificadas.
Mutuaciones
una mutación es un cambio en la información genética (genotipo) de un ser vivo (muchas veces por contacto con mutágenos), que produce una variación en las características de este que se presenta de manera espontánea y súbita, que se puede heredar a la descendencia. Este cambio estará presente en una pequeña proporción de la población (variante) o del organismo (mutación). La unidad genética capaz de mutar es el gen, la unidad de información hereditaria que forma parte del ADN.
En los seres multicelulares, las mutaciones solo pueden ser heredadas cuando afectan a las células reproductivas. Una consecuencia de las mutaciones puede ser, por ejemplo, una enfermedad genética. Sin embargo, aunque a corto plazo pueden parecer perjudiciales, las mutaciones son esenciales para nuestra existencia a largo plazo. Sin mutación no habría cambio, y sin cambio la vida no podría evolucionar.
Definición
La definición que en su obra de 1901 "La teoría de la mutación" Hugo de Vries dio de la mutación (del latín mutare = cambiar) era la de cualquier cambio heredable en el material hereditario que no se puede explicar mediante segregación o recombinación. Más tarde se descubrió que lo que De Vries llamó mutación en realidad eran más bien recombinaciones entre genes.
La definición de mutación a partir del conocimiento de que el material hereditario es el ADN y de la propuesta de la doble hélice para explicar la estructura del material hereditario (Watson y Crick,1953), sería que una mutación es cualquier cambio en la secuencia de nucleótidos del ADN. Cuando dicha mutación afecta a un solo gen, se denomina mutación génica. Cuando es la estructura de uno o varios cromosomas lo que se ve afectado, mutación cromosómica. Y cuando una o varias mutaciones provocan alteraciones en todo el genoma se denominan, mutaciones genómicas.
Mutación somática y mutación en la línea germinal
Mutación somática: es la que afecta a las células somáticas del individuo. Como consecuencia aparecen individuos mosaico que poseen dos líneas celulares diferentes con distinto genotipo. Una vez que una célula sufre una mutación, todas las células que derivan de ella por divisiones mitóticas heredarán la mutación (herencia celular). Un individuo mosaico originado por una mutación somática posee un grupo de células con un genotipo diferente al resto, cuanto antes se haya dado la mutación en el desarrollo del individuo mayor será la proporción de células con distinto genotipo. En el supuesto de que la mutación se hubiera dado después de la primera división del cigoto (en estado de dos células), la mitad de las células del individuo adulto tendrían un genotipo y la otra mitad otro distinto. Las mutaciones que afectan solamente a las células de la línea somática no se transmiten a la siguiente generación.23
Mutaciones en la línea germinal: son las que afectan a las células productoras de gametos apareciendo, de este modo, gametos con mutaciones. Estas mutaciones se transmiten a la siguiente generación y tienen una mayor importancia en la evolución biológica.
Tipos de mutación según sus consecuencias
Las consecuencias fenotípicas de las mutaciones son muy variadas, desde grandes cambios hasta pequeñas diferencias tan sutiles que es necesario emplear técnicas muy desarrolladas para su detección.
Mutaciones morfológicas
Afectan a la morfología del individuo, a su distribución corporal. Modifican el color o la forma de cualquier órgano de un animal o de una planta. Suelen producir malformaciones. Un ejemplo de una mutación que produce malformaciones en humanos es aquella que determina la neurofibromatosis. Esta es una enfermedad hereditaria, relativamente frecuente (1 en 3.000 individuos), producida por una mutación en el cromosoma 17 y que tiene una penetrancia del 100 % yexpresividad variable. Sus manifestaciones principales son la presencia de neurofibromas, glioma del nervio óptico, manchas cutáneas de color café con leche, hamartomas del iris, alteraciones óseas (displasia del esfenoide, adelgazamiento de la cortical de huesos largos). Con frecuencia hay retardo mental y macrocefalia.4
Mutaciones letales y deletéreas
Son las que afectan la supervivencia de los individuos, ocasionándoles la muerte antes de alcanzar la madurez sexual. Cuando la mutación no produce la muerte, sino una disminución de la capacidad del individuo para sobrevivir y/o reproducirse, se dice que la mutación es deletérea. Este tipo de mutaciones suelen producirse por cambios inesperados en genes que son esenciales o imprescindibles para la supervivencia del individuo. En general las mutaciones letales son recesivas, es decir, se manifiestan solamente en homocigosis o bien, en hemicigosis para aquellos genes ligados alcromosoma X en humanos, por ejemplo.25
Mutaciones condicionales
Las mutaciones condicionales (incluidas las condicionalmente letales) son muy útiles para estudiar aquellos genes esenciales para la bacteria. En estos mutantes hay que distinguir dos tipos de condiciones:
condiciones restrictivas (también llamadas no-permisivas): son aquellas condiciones ambientales bajo las cuales el individuo pierde la viabilidad, o su fenotipo se ve alterado, debido a que el producto afectado por la mutación pierde su actividad biológica.
condiciones permisivas: son aquellas bajo las cuales el producto del gen mutado es aún funcional.
Mutaciones bioquímicas o nutritivas
Son los cambios que generan una pérdida o un cambio de alguna función bioquímica como, por ejemplo, la actividad de una determinadaenzima. Se detectan ya que el organismo que presenta esta mutación no puede crecer o proliferar en un medio de cultivo por ejemplo, a no ser que se le suministre un compuesto determinado. Los microorganismos constituyen un material de elección para estudiar este tipo de mutaciones ya que las cepas silvestres solo necesitan para crecer un medio compuesto por sales inorgánicas y una fuente de energía como laglucosa. Ese tipo de medio se denominamínimoy las cepas que crecen en él se dicenprototróficas. Cualquier cepa mutante para un gen que produce una enzima perteneciente a unavía metabólicadeterminada, requerirá que se suplemente el medio de cultivo mínimo con el producto final de la vía o ruta metabólica que se encuentra alterada. Esa cepa se llamaauxotróficay presenta una mutación bioquímica o nutritiva.
Mutaciones de pérdida de función
Las mutaciones suelen determinar que la función del gen en cuestión no se pueda llevar a cabo correctamente, por lo que desaparece alguna función del organismo que la presenta. Este tipo de mutaciones, las que suelen ser recesivas, se denominan mutaciones de pérdida de función. Un ejemplo es la mutación del gen hTPH2 que produce la enzimatriptófano hidroxilasa en humanos. Esta enzima está involucrada en la producción de serotonina en el cerebro. Una mutación (G1463A) de hTPH2 determina aproximadamente un 80 % de pérdida de función de la enzima, lo que se traduce en una disminución en la producción de serotonina y se manifiesta en un tipo de depresión llamada depresión unipolar.7
Mutaciones de ganancia de función
Cuando ocurre un cambio en el ADN, lo más normal es que corrompa algún proceso normal del ser vivo. Sin embargo, existen raras ocasiones donde una mutación puede producir una nueva función al gen, generando un fenotipo nuevo. Si ese gen mantiene la función original, o si se trata de un gen duplicado, puede dar lugar a un primer paso en la evolución. Un caso es la resistencia a antibióticos desarrollada por algunas bacterias (por eso no es recomendable hacer un uso abusivo de algunos antibióticos ya que finalmente el organismo patógeno irá evolucionando y el antibiótico no le hará ningún efecto).
Tipos de mutación según el mecanismo causal
Según el mecanismo que ha provocado el cambio en el material genético, se suele hablar de tres tipos de mutaciones:mutaciones cariotípicas o genómicas, mutaciones cromosómicas y mutaciones génicas o moleculares. En el siguiente cuadro se describen los diferentes tipos de mutaciones y los mecanismos causales de cada una de ellas.23
Hay una tendencia actual a considerar como mutaciones en sentido estricto solamente las génicas, mientras que los otros tipos entrarían en el término de aberraciones cromosómicas.
Mutuaciones Cromosómicas
Definición
Las mutaciones cromosómicas son modificaciones en el número total de cromosomas, la duplicación o supresión de genes o de segmentos de un cromosoma y la reordenación del material genético dentro o entre cromosomas. Pueden ser vistas al microscopio, sometiendo a los cromosomas a la “técnica de bandas”. De esta manera se podrá confeccionar el cariotipo.
Introducción
Las alteraciones de la dotación diploide de cromosomas se denominan aberraciones cromosómicas o mutaciones cromosómicas.
Los lugares frágiles son constricciones o brechas que aparecen en regiones particulares de los cromosomas con una predisposición a romperse en determinadas condiciones.
El estudio de las series normales y anormales de cromosomas se conoce como citogenética.
Aneuploidía
La alteración en el número de cromosomas es denominada aneuploidía. La aneuploidía se define como la pérdida o ganancia de cromosomas completos en un individuo. Este fenómeno puede ocurrir en cualquiera de los cromosomas autosómicos (del 1 al 22) o sexuales (X e Y).
La ganancia de un cromosoma completo en una célula es denominadatrisomía(2n+1), y en ese caso el cariotipo del individuo estaría formado por 47 cromosomas. Probablemente la trisomía más conocida sea el Síndrome de Down (trisomía del cromosoma 21). La pérdida de un cromosoma es denominada monosomía(2n-1) y el número de cromosomas de cada célula sería 45. La única monosomía viable en el hombre es la del cromosoma X, que origina en los individuos que la padecen el Síndrome de Turner.
En las células somáticas hay un mecanismo que inactiva a todos los cromosomas X menos uno, la ganancia o perdida de un cromosoma sexual en genoma diploide altera el fenotipo normal, dando lugar a los síndromes deKlinefelter o de Turner, respectivamente.
Tal variación cromosómica se origina como un error aleatorio durante la producción de gametos. La no disyunción es el fallo de los cromosomas o de las cromatidas en separarse y desplazarse a los polos opuestos en la meiosis. Cuando esto ocurre se desbarata la distribución normal de los cromosomas en los gametos. El cromosoma afectado puede dar lugar a gametos anormales con dos miembros o con ninguno. La fecundación de estos con un gameto haploide normal da lugar a zigotos con tres miembros (trisomía) o con solo uno (monosomía) de este cromosoma. La no disyunción da lugar a una serie de situaciones aneuploides autosómicas en la especie humana y en otros organismos.
Síndrome de Klinefelter
El síndrome de Klinefelter se considera la anomalía gonosómica más común en los humanos. Los afectados presentan un cromosoma “X” supernumerario lo que conduce a fallo testicular primario con infertilidad e hipoandrogenismo. A pesar de la relativa frecuencia del padecimiento en recién nacidos vivos, se estima que la mitad de los productos 47, XXY se abortan de manera espontánea.
Síndrome de Turner
El síndrome de Turner o Monosomía X es una enfermedad genética caracterizada por presencia de un solo 'cromosoma X'. La falta de cromosoma Y determina el sexo femenino de todos los individuos afectados, y la ausencia de todo o parte del segundo cromosoma X determina la falta de desarrollo de los caracteres sexuales primarios y secundarios. Esto confiere a las mujeres que padecen el síndrome de Turner un aspecto infantil e infertilidad de por vida.
Variaciones en estructura y ordenación de los cromosomas
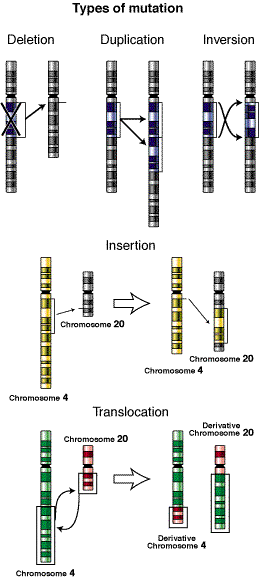
El otro tipo de aberración cromosómica incluye cambios estructurales que eliminan, añaden o reordenan partes sustanciales de uno o más cromosomas, se encuentran las deleciones y las duplicaciones de genes o de parte de un cromosoma y las reordenaciones del material genético mediante las que segmentos de un cromosoma se invierten, se intercambian con un segmento de un cromosoma no homólogo o simplemente se transfieren a otro cromosoma. Los intercambios y las transferencias se denominan translocaciones, en las que la localización de un gen esta cambiada dentro del genoma.
Estos cambios estructurales se deben a una o más roturas distribuidas a lo largo del cromosoma, seguidas por la pérdida o la reordenación del material genético. Los cromosomas pueden romperse espontáneamente, pero la tasa de roturas puede aumentar en células expuestas a sustancias químicas o a radiación. Aunque los extremos normales de los cromosomas, los telómeros, no se fusionan fácilmente con extremos nuevos de cromosomas rotos o con otros telómeros, los extremos producidos en los puntos de rotura son cohesivos (“pegajosos”) y pueden reunirse con otros extremos rotos. Si la rotura y reunión no restablece las relaciones originales y si la alteración se produce en el plasma germinal, los gametos tendrán una reordenación estructural que será heredable.
Si la aberración se encuentra en un homólogo, pero no en el otro, se dice que los individuos son heterocigotos para la aberración. En tales casos se producen configuraciones raras en el apareamiento durante la sinapsis meiótica.
Si no hay pérdida o ganancia de material genético, los individuos que llevan la aberración en heterocigosis en uno de los dos homólogos probablemente no quedaran afectados en su fenotipo. Los complicados apareamientos de las ordenaciones dan lugar a menudo a gametos con duplicaciones o deficiencias de algunas regiones cromosómicas. Cuando esto ocurre, los descendientes de “portadores” de ciertas aberraciones tienen a menudo una mayor probabilidad de presentar cambios fenotípicos.
Translocaciones
Las translocaciones ocurren cuando un fragmento de ADN es transferido desde un cromosoma a otro no homólogo. Se incluyen:
Traslocaciones recíprocas: Es una translocación balanceada. No hay pérdida o ganancia neta de material genético. Los individuos portadores de translocaciones recíprocas no suelen presentar ningún fenotipo. Sin embargo, estos individuos portadores tienen riesgo de producir descendencia con translocaciones desbalanceadas, que sí pueden estar asociadas a patologías o conducir al aborto del feto. También pueden ser un problema las translocaciones recíprocas de novo, si la rotura del cromosoma tiene lugar en genes importantes.
Translocaciones desbalanceadas: Hay pérdida o ganancia de material genético respecto al genotipo silvestre. Pueden suponer un problema para el individuo portador de las mismas.
Translocacion Robertsoniana: Son translocaciones "casi" equilibradas. Este tipo de mutación cromosómica tiene lugar con la fusión de los brazos largos de dos cromosomas acrocéntricos. Los brazos cortos de ambos cromosomas se pierden. El cariotipo de los individuos con este tipo de translocaciones muestra 45 cromosomas, sin embargo no produce anomalía fenotípicas puesto que casi todo el material genético está presente. Las translocaciones Robertsonianas en un individuo pueden ser responsables de translocaciones desbalanceadas en su descendencia.
Las deleciones pueden eliminar o inactivar los genes que controlan el ciclo celular;
Las inversiones y las translocaciones pueden causar rupturas en los genes supresores de tumores, fusionar genes que producen proteínas cancerígenas o mover genes a nuevas ubicaciones, donde quedan bajo la influencia de diferentes secuencias reguladoras.
El papel de las mutaciones en el cáncer.
Las mutaciones en los genes regulatorios claves (los supresores de tumor y los protooncogenes) alteran el estado de las células y pueden causar el crecimiento irregular visto en el cáncer. Para casi todos los tipos de cáncer que se han estudiado hasta la fecha, parece que la transición de una célula sana y normal a una célula cancerosa es una progresión por pasos que requiere cambios genéticos en varios oncogenes y supresores de tumor diferentes. Esta es la razón por la cual el cáncer es mucho más prevalente en individuos de edades mayores. Para generar una célula cancerosa, una series de mutaciones deben ocurrir en la misma célula. Ya que la probabilidad de que cualquier gen sea mutado es muy baja, es razonable decir que la probabilidad de varias mutaciones en la misma célula es aún más improbable.
Mutaciones genómicas o numéricas
La trisomía en el par cromosómico 21 en los humanos ocasiona el Síndrome de Down
Son las mutaciones que afectan al número de cromosomas o todo el complemento cromosómico (todo el genoma).
Poliploidía: Es la mutación que consiste en el aumento del número normal de “juegos de cromosomas” . Los seres poliploides pueden ser autopoliploides, si todos los juegos proceden de la misma especie, o alopoliploides, si proceden de la hibridación, es decir, del cruce de dos especies diferentes.
Haploidía: Son las mutaciones que provocan una disminución en el número de juegos de cromosomas.
Aneuploidía: Son las mutaciones que afectan solo a un número de ejemplares de un cromosoma o más, pero sin llegar a afectar al juego completo. Las aneuploidías pueden ser monosomías, trisomías, tetrasomías, etc, cuando en lugar de dos ejemplares de cada tipo de cromosomas, que es lo normal, hay o solo uno, o tres, o cuatro, etc. Entre las aneuplodías podemos encontrar diferentes tipos de trastornos genéticos en humanos como pueden ser:
Son las mutaciones que alteran la secuencia de nucleótidos del ADN. Estas mutaciones pueden llevar a la sustitución de aminoácidos en las proteínas resultantes (se denominan mutaciones no sinónimas). Un cambio en un solo aminoácido puede no ser importante si es conservativo y ocurre fuera del sitio activo de la proteína. Así, existen las denominadasmutaciones sinónimas o "mutaciones silenciosas" en las que la mutación altera la base situada en la tercera posición delcodón pero no causa sustitución aminoacídica debido a la redundancia del código genético. El aminoácido insertado será el mismo que antes de la mutación. También, en el caso de las mutaciones neutras, el aminoácido insertado es distinto pero con unas propiedades fisicoquímicas similares, por ejemplo la sustitución de glutámico por aspártico puede no tener efectos funcionales en la proteína debido a que los dos son ácidos y similares en tamaño. También podrían considerarse neutras aquellas mutaciones que afecten a zonas del genoma sin función aparente, como las repeticiones en tándem o dispersas, las zonas intergénicas y los intrones.8
De lo contrario, la mutación génica o también llamada puntual, puede tener consecuencias severas, como por ejemplo:
La sustitución de valina por ácido glutámico en la posición 6 de la cadena polipeptídica de la beta-globina da lugar a la enfermedad anemia falciforme en individuos homocigóticos debido a que la cadena modificada tiene tendencia a cristalizar a bajas concentraciones de oxígeno.
Las proteínas del colágeno constituyen una familia de moléculas estructuralmente relacionadas que son vitales para la integridad de muchos tejidos incluidos la piel y los huesos. La molécula madura del colágeno está compuesta por 3 cadenas
polipeptídicas unidas en una triple hélice. Las cadenas se asocian primero por su extrempo C-terminal y luego se enroscan hacia el extremo N-terminal. Para lograr este plegado, las cadenas de colágeno tienen una estructura repetitiva de 3 aminoácidos: glicina - X - Y (X es generalmente prolina y Y puede ser cualquiera de un gran rango de aminoácidos). Unamutación puntual que cambie un solo aminoácido puede distorsionar la asociación de las cadenas por su extremo C-terminal evitando la formación de la triple hélice, lo que puede tener consecuencias severas. Una cadena mutante puede evitar la formación de la triple hélice, aun cuando haya 2 monómeros de tipo salvaje. Al no tratarse de una enzima, la pequeña cantidad de colágeno funcional producido no puede ser regulada. La consecuencia puede ser la condición dominante letal osteogénesis imperfecta.
Bases moleculares de la mutación génica
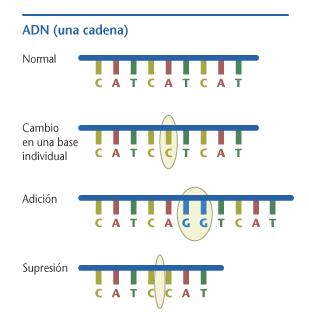
Mutación por sustitución de bases: Se producen al cambiar en una posición un par de bases por otro (son las bases nitrogenadas las que distinguen los nucleótidos de una cadena). Distinguimos dos tipos que se producen por diferentes mecanismos bioquímicos:8
Mutaciones transicionales o simplemente transiciones, cuando un par de bases es sustituido por su alternativa del mismo tipo. Las dos bases púricas son adenina (A) y guanina (G), y las dos pirimídicas son citosina (C) y timina (T). La sustitución de un par AT, por ejemplo, por un par GC, sería una transición.
Mutaciones transversionales o transversiones, cuando un par de bases es sustituida por otra del otro tipo. Por ejemplo, la sustitución del par AT por TA o por CG.
Mutaciones de corrimiento estructural, cuando se añaden o se quitan pares de nucleótidos alterándose la longitud de la cadena. Si se añaden o quitan pares en un número que no sea múltiplo de tres (es decir si no se trata de un número exacto de codones), las consecuencias son especialmente graves, porque a partir de ese punto, y no solo en él, toda la información queda alterada. Hay dos casos:
Mutación por pérdida o deleción de nucleótidos: en la secuencia de nucleótidos se pierde uno y la cadena se acorta en una unidad.
Mutación por inserción de nuevos nucleótidos: Dentro de la secuencia del ADN se introducen nucleótidos adicionales, interpuestos entre los que ya había, alargándose correspondientemente la cadena.8
Las mutaciones de corrimiento del marco de lectura también pueden surgir por mutaciones que interfieren con el ayuste del ARN mensajero. El comienzo y final de cada intrón en un gen están definidos por secuencias conservadas de ADN. Si un nucleótido muta en una de las posiciones altamente conservada, el sitio no funcionará más, con las consecuencias predecibles para el ARNm maduro y la proteína codificada. Hay muchos ejemplos de estas mutaciones, por ejemplo, algunas mutaciones en el gen de la beta globina en la beta talasemia son causadas por mutaciones de los sitios de ayuste.
Mutaciones espontáneas o inducidas
Las mutaciones pueden ser espontáneas o inducidas. Las primeras son aquellas que surgen normalmente como consecuencia de errores durante el proceso de replicación del ADN. Tales errores ocurren con una probabilidad de 10−7en células haploides y 10−14 en diploides.8
Mutaciones inducidas
Las mutaciones inducidas surgen como consecuencia de la exposición a mutágenos químicos o biológicos o a radiaciones. Entre los mutágenos químicos se pueden citar:
los análogos de bases del ADN (como la 2-aminopurina), moléculas que se parecen estructuralmente a las bases púricas o pirimidínicas pero que muestran propiedades de apareamiento erróneas;
los agentes alquilantes como la nitrosoguanidina, que reacciona directamente con el ADN originando cambios químicos en una u otra base y produciendo también apareamientos erróneos;
y, por último, los agentes intercalantes como las acridinas, que se intercalan entre 2 pares de bases del ADN, separándolas entre sí.
Como mutágenos biológicos podemos considerar la existencia de transposones o virus capaces de integrarse en el genoma.
Las radiaciones ionizantes (rayos X, rayos cósmicos y rayos gamma) y no ionizantes (sobre todo la radiación ultravioleta) también inducen mutaciones en el ADN; las primeras se originan por los radicales libres que reaccionan con el ADN inactivándolo, y las segundas aparecen como consecuencia de la formación de dímeros de pirimidina en el ADN, es decir, como consecuencia de la unión covalente de 2 bases pirimidínicas adyacentes.
Un agente utilizado a menudo para inducir mutaciones (mutagénesis) en organismos experimentales es el EMS (sulfato de etilmetano). Este mutágeno puede alterar la secuencia del DNA de diversas maneras como modificar químicamente las bases de G en DNA. Esta alteración en la secuencia de un gen se conoce como mutación puntual.
Mutaciones espontáneas
Las principales causas de las mutaciones que se producen de forma natural o normal en las poblaciones son tres: los errores durante la replicación del ADN, las lesiones o daños fortuitos en el ADN y la movilización en el genoma de los elementos genéticos transponibles.
Errores en la replicación
Durante la replicación del ADN pueden ocurrir diversos tipos de errores que conducen a la generación de mutaciones. Los tres tipos de errores más frecuentes son:
La tautomería: las bases nitrogenadas se encuentran habitualmente en su forma cetónica y con menos frecuencia aparecen en su forma tautomérica enólica o imino. Las formas tautoméricas o enólicas de las bases nitrogenadas (A*, T*, G* y C*) muestran relaciones de apareamiento distintas que las formas cetónicas: A*-C, T*-G, G*-T y C*-A. El cambio de la forma normal cetónica a la forma enólica produce transiciones. Los errores en el apareamiento incorrecto de las bases nitrogenadas pueden ser detectados por la función correctora de pruebas de la ADN polimerasa III.
Las mutaciones de cambio de fase o pauta de lectura: se trata de inserciones o deleciones de uno o muy pocos nucleótidos. Según un modelo propuesto por Streisinger, estas mutaciones se producen con frecuencia en regiones con secuencias repetidas. En las regiones con secuencias repetidas, por ejemplo, TTTTTTTTTT..., o por ejemplo, GCGCGCGCGCGCG...., durante la replicación se puede producir el deslizamiento de una de las dos hélices (la hélice molde o la de nueva síntesis) dando lugar a lo que se llama "apareamiento erróneo deslizado". El deslizamiento de la hélice de nueva síntesis da lugar a una adición, mientras que el deslizamiento de la hélice molde origina una deleción. En el gen lac I (gen estructural de la proteína represora) de E. coli se han encontrado puntos calientes (regiones en las que la mutación es muy frecuente) que coinciden con secuencias repetidas: un ejemplo es el punto caliente CTGG CTGG CTGG.
Deleciones y duplicaciones grandes: las deleciones y duplicaciones de regiones relativamente grandes también se han detectado con bastante frecuencia en regiones con secuencias repetidas. En el gen lac I de E. coli se han detectado deleciones grandes que tienen lugar entre secuencias repetidas. Se cree que estas mutaciones podrían producirse por un sistema semejante al propuesto por Streisinger ("Apareamiento erróneo deslizado") o bien por entrecruzamiento desigual..






{kind=link}